比较转录组分析莲WRKY基因家族响应激素SA和JA的表达谱毕业论文
2020-02-19 14:35:02
摘 要
WRKY转录因子在植物生长发育的各个阶段发挥着重要的作用。本研究通过比较转录组分析莲WRKY基因家族响应激素水杨酸(SA)和茉莉酸(JA)的表达谱。在莲中我们发现了60个候选WRKY基因,其中13个基因能响应SA,19个基因响应JA,其中7个基因WRKY4a、WRKY7b、WRKY28a、WRKY31a、WRKY31b、WRKY33a和WRKY65b可被两种激素均诱导且表达水平增加,推测SA和JA介导的信号途径在莲中的拮抗关系并不显著。通过比较SA和JA诱导WRKY基因的模式不同,发现了SA的诱导是瞬时的,但JA的诱导则是需要一定的时间才能进行。本研究为下一步研究WRKY基因的功能提供了必要的理论基础。
关键词:莲;WRKY;水杨酸;茉莉酸;转录组分析
Abstract
WRKY transcription factor plays an important role in various stages of plant growth and development. In this study, the expression profiles of lotus WRKY genes under the salicylic acid (SA) and jasmonic acid (JA) treatment were analyzed by using transcriptome data. In lotus, we found 60 candidate WRKY genes, 13 of which could respond to SA, and 19 could respond to JA, and 7 of which, WRKY4a, WRKY7b, WRKY28a, WRKY31a, WRKY31b, WRKY33a and WRKY65b, could be induced by both and their expression levels increased, suggesting that SA and JA pathways are mutually promoting in lotus. By comparing the patterns of SA and ja-induced WRKY genes, it was found that the induction of SA was instantaneous, while JA induction took some time. This study provides a necessary theoretical basis for the further study on the function of WRKY gene.
Key Word:Lotus;WRKY;salicylic acid;jasmonic acid;transcriptome analysis
目 录
第1章 绪论 1
1.1 WRKY转录因子概述 1
1.1.1 WRKY转录因子简介 1
1.1.2 WRKY转录因子的结构及分类 1
1.1.3 WRKY转录因子的作用 2
1.2 WRKY转录因子与植物激素的相互作用 5
1.3莲中WRKY转录因子简介 5
第2章 比较转录组分析莲WRKY基因家族响应激素SA和JA的表达谱 7
2.1研究内容与方法 7
2.1.1 内容 7
2.1.2方法 7
2.2结果与分析 9
2.2.1原始测序数据总览 9
2.2.2基因表达量分析 10
2.2.3差异基因分析 14
2.3讨论 17
第3章 结论 20
参考文献 21
附录A 24
致谢 30
第1章 绪论
1.1 WRKY转录因子概述
1.1.1 WRKY转录因子简介
真核生物的转录过程可以分为四个阶段:模板识别、转录起始、转录延伸和转录的终止。转录起始比较复杂,需要多种复合物同时参与[1]。研究表明除了RNA聚合酶之外,真核生物的转录的起始至少还需要7种蛋白辅助因子参与,这些辅助因子被称为转录因子(transcription factor, TF)[2]。转录因子从蛋白结构上分析可以被分为四个区域;DNA结合区、转录调控区、寡聚化位点和核定位信号。转录因子通过这些功能结构和启动子的顺式元件作用或者同其他的转录因子的功能结构相互作用进而调控目标基因的转录过程[3]。WRKY转录因子是植物中最大的转录调控因子家族之一,在调控植物生长发育以及植物抗生物及非生物胁迫过程中起着重要作用[4, 5]。研究表明,WRKY蛋白在植物生命活动中扮演着激活和抑制因子的角色。绿藻和陆地植物的整个谱系中都发现了WRKY转录家族,并且随着物种的进化,该家族成员正逐渐扩大[6]。自从第一个WRKY家族基因SPF1从甘薯中被发现以来[7, 8],前人对WRKY蛋白的研究取得了重大进展,并且对于WRKY转录因子的研究也不限于拟南芥等模式植物[9]。
1.1.2 WRKY转录因子的结构及分类
WRKY转录因子因具有高度保守的七肽结构WRKYGQK(少数蛋白基序为WRKYGKK、WKKYGQK、WRKYGQR、WRKYGEK等)而得名。常见的WRKY结构域长度约为60个氨基酸残基[9],除了N端含有WRKY的特征氨基酸序列外,在C端还存在一个非典型的特殊锌指结构,该结构可分为两类,分别为CX4-5CX22-23HXH或CX7CX23HXC[10]。WRKY结构域由4个片状β折叠构成,锌配位的Cys/His残基会形成一个锌结合袋,与锌结合。WRKYGQK残基对应于大多数N端β链,其表面存在着部分凸起,从而允许主要的DNA沟槽进入并与DNA接触[9]。W盒是WRKY蛋白的特异DNA结合区,大多数WRKY基因的启动子区域存在同向排列或形成回文结构的W盒,这与WRKY基因的调控方式有关[11]。推测WRKY基因通过控制下游基因顺式元件W盒的特异结合从而调控下游基因的表达[12, 13]。除了 WRKY结构域和锌指结构之外,WRKY 蛋白还其他的特殊结构,比如核定位信号、亮氨酸拉链结构、激酶域、TIR-NBS-LRR 以及富含谷氨酸酰胺域、丝氨酸/苏氨酸域和脯氨酸域等,这些特殊结构使得WRKY转录因子拥有不同的转录调控功能[13]。
根据WRKY结构域的个数和锌指结构的特征,WRKY转录因子可分为三类,如图1.1所示。第I类WRKY转录因子含有两个WRKY结构域;而第II和第III类家族成员只有一个WRKY结构域。另外,第I和第II类成员所具有的锌指结构为C2 -H2(C-X 4-5 -CX 22-23 -H-X1 -H);而第III类WRKY蛋白所含的锌指结构则为C2 -HC(C-X 7 -C-X 23 -H-X-C),其中X可以是任何氨基酸[4]。依据WRKY保守域外的氨基酸序列与其种族发生的间距,第II类又可分为5个亚族, IIa-IIe [14]。
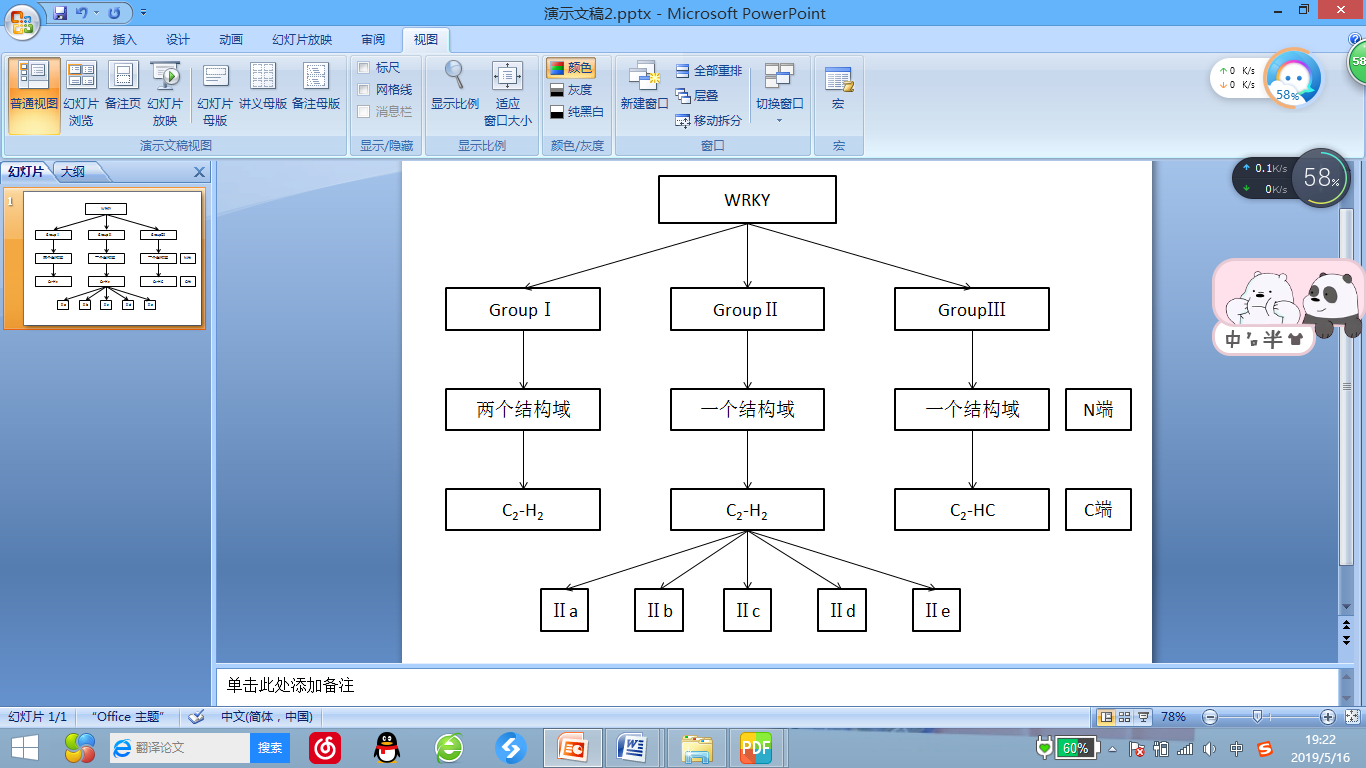
图1.1 WRKY转录因子的分类
1.1.3 WRKY转录因子的作用
1.1.3.1 WRKY转录因子与植物生长发育
拟南芥AtWRKY10基因,又被称为MINISEED3,表达于花粉和球状胚中,也表达于从双核到细胞化阶段的胚乳发育中。这种基因敲除突变的拟南芥纯合子将会产生迷你种子。这些种子的发育表现为胚的发育减少和胚乳的早期细胞化,这是由于AtWRKY10的缺失影响胚的发育[9, 15]。在水稻中,OsWRKY78基因敲除突变体导致种子体积变小,说明OsWRKY78在种子发育调控中发挥着正向调控作用。
植物的衰老是植物生长发育的重要一环,在植物衰老的过程中,许多WRKY转录因子都被证明参与了反应。通过分析拟南芥WRKY基因的表达谱,发现WRKY转录因子参与了植物的衰老过程。AtWRKY6的表达在植物衰老的过程中明显上调。其他调节衰老的WRKY基因还有AtWRKY53和AtWRKY70等。其中,AtWRKY53的过表达导致植物衰老;AtWRKY70基因敲除的株系表明其是衰老的负调控因子。这同样说明了WRKY转录因子家族成员对衰老这一过程既存在正向调节作用,也有负向调节作用[9]。
1.1.3.2 WRKY转录因子与生物胁迫
当植物受到生物胁迫时,WRKY 转录因子会激活水杨酸(SA)、茉莉酸(JA)和乙烯(ET)等介导的信号通路,然后改变与之相关基因的转录水平[16]。WRKY转录因子是植物天然免疫反应过程中的核心组成部分,这些免疫反应包括病原体相关分子模式触发的免疫反应(PAMP-triggered immunity,PTI)和效应因子二次免疫反应(Effector-triggered immunity, ETI)。研究表明,WRKY基因的过度表达或下调对植物防御有影响。对拟南芥WRKY11和WRKY70两个转录因子的研究发现,二者在蜡样芽孢杆菌诱发的系统抗性中发挥着重要的调控作用。调控的基本途径是WRKY11基因能够激活茉莉酸信号途径,WRKY70则是激活水杨酸信号途径,从而共同调节了蜡样芽孢杆菌诱发的系统抗性这一过程[17]。另有研究发现,水稻中OsWRKY45-1 和OsWRKY45-2,协同正调控水稻对稻瘟病菌的抗性。然而,这两个转录因子却对水稻白叶枯病菌和细菌性条斑病菌表现出相反的调控作用。进一步的研究显示,OsWRKY45-1同时调节了水杨酸和茉莉酸的水平,但OsWRKY45-2似乎只对茉莉酸水平有显著的调节作用,对水杨酸影响不大[18]。我们由此可以推测,不同的基因可能是通过不同的途径对植物生理活动进行调控。同样,在抵抗水稻稻瘟病方面, OsWRKY42通过抑制与茉莉酸途径相关基因,从而负调控稻瘟病。此外,水稻 OsWRKY4通过正调控茉莉酸和乙烯途径来提高其对水稻纹枯病的抗性,而OsWRKY51也被证实其过量表达可提高水稻对白叶枯病的抗性[17]。
WRKY基因除了参与植物抗性的调节,还可以通过多种途径减少植物被草食动物侵害[19]。烟草中NtWRKY3和NtWRKY6能够协同调控烟草对草食动物的反应。NtWRKY3是六重树幼虫口腔分泌物中的脂肪酸-氨基偶联物诱导NtWRKY6所必需的基因,如果使其中一个或两个基因沉默,食草动物攻击植物的几率会增加。WRKY转录因子同样也被证实参与了植物关于线虫的抗性反应,拟南芥基因AtWRKY23在线虫感染后几乎立即上调,而敲除该基因,敲除株系相比较野生型则显示出对囊肿线虫异源性的抗性增强[9]。有研究表明,水稻OsWRKY45 能被机械损伤、褐飞虱取食及外源茉莉酸和水杨酸等多种因素诱导表达,如果通过某种途径降低OsWRKY45基因的表达,就可以增加植物体内的过氧化氢和乙烯含量,从而减少植物对褐飞虱的养分供给,然后降低其产卵量,延迟若虫的生长发育,最终达到降低种群密度的效果[17]。
1.1.3.3 WRKY转录因子与非生物胁迫
WRKY转录因子在植物非生物胁迫应答反应中同样起着重要的作用。植物响应非生物胁迫的具体模式是:当植物感受非生物胁迫信号,例如干旱、盐胁迫、冷胁迫和热胁迫等等时,植物会迅速将信号传递到细胞的内部,植物细胞会做出一系列反应,比如改变细胞内部Ca2 离子、环腺苷酸(cAMP)或活性氧自由基等浓度,进而激活MPK和CDPK等蛋白激酶,调控相关转录因子(AP2/ERFBP, MYB, WRKY, NAC和bZIP等)活性,最后产生非生物胁迫反应[18]。
干旱是限制植物正常生长的关键因素之一,土壤干旱时,植物生长缓慢或者几乎完全不能生长,WRKY转录家族中的许多成员都有被发现响应干旱胁迫。干旱因为水分的减少经常会引起土壤盐渍化,所以植物在受到干旱胁迫的同时往往也会受到盐胁迫。拟南芥双突变体wrky25wrky33对盐浓度变化具有高度敏感性,WRKY25和WRKY33的过表达都会使植物获得盐胁迫耐受性,从而适应环境变化[4]。过表达AtWRKY57基因也能增加拟南芥的耐旱性,其原理是AtWRKY57蛋白能够与RD29A和NCED3基因的启动子中的W盒结合,促进RD29A和NCED3的表达,从而增强拟南芥的耐旱和耐盐性。当OsWRKY45 和OsWRKY72基因过表达时,水稻对干旱和盐胁迫的耐受性也大大增加。另外,拟南芥 wrky63(abo3)突变体能够改变植物对ABA(脱落酸)的敏感性和耐旱性,进一步的分析发现, AtWRKY63位于ABA 信号传导途径中 ABI1、ABI2和ABI5的下游,ABF2、RD29A和COR4 的上游,所以拟南芥对脱落酸的敏感性和耐旱性会发生变化[5]。
热胁迫一般被认为是主要的非生物胁迫因素,在较高的温度下,植物无法正常生长,农作物若是在较高温度下生长,收成将会受到严重影响。植物在受到高温危害后,经常会产生各种病害特征,例如树干开裂等等[20]。拟南芥的AtWRKY39转录因子是第 II 类 WRKY 蛋白家族的成员,通常会对多种胁迫作出反应。有实验表明,AtWRKY39蛋白是植物热激反应的正向调控因子,AtWRKY39过表达可使植物的耐热性增强。热处理诱导AtWRKY39 基因表达量增加的同时,水杨酸和茉莉酸信号途径正向共调节AtWRKY39的表达。与之相关的还有,当小麦处于高温环境时,TaWRKY70 的转录物明显增加。当植物受到乙烯、水杨酸和冷胁迫时,TaWRKY70的表达水平上升;受到茉莉酸甲酯和热胁迫时,表达量下降。此外,TaWRKY33转基因株系的耐热性也大大增强[3]。
植物的生长过程中,在炎热的夏天会受到热胁迫,在寒冷的冬天则会受到冷胁迫。植物处在低温条件下时,WRKY转录因子也发挥着重要的作用,通过调节多种途径使植物适应寒冷环境。水稻的一对等位基因即OsWRKY45-1和OsWRKY45-2,两者对低温环境有着相同的响应,增强植物对低温的抗性 [6]。
1.2 WRKY转录因子与植物激素的相互作用
植物激素(phytohormone)是一种植物生长物质,能够调节植物生长发育。植物激素是一类在植物体内生成,从产生的地方运送到其他地方,对植物生长发育有显著影响的微量有机物[20]。常见的植物激素有生长素、细胞分裂素、乙烯、水杨酸、茉莉酸等。水杨酸、茉莉酸和乙烯作为植物防御调节的主要信号,对植物响应环境有重要意义[21]。
水杨酸(salicylic,SA),是邻羟基苯甲酸的桂皮酸的衍生物,是植物体内重要的信号分子。SA在植物抗病的过程扮演者重要的角色,部分植物在被病原微生物侵染之后,SA会开始合成,然后产生相应的蛋白质,提高植物的抗性。SA还有其他的生理功能,比如诱导浮萍开花等[20]。茉莉酸(jasmonic acid, JA)在植物抵抗昆虫的过程中充当信号分子,JA能够诱导产生的蛋白质有十几种,大多是与植物抵抗病虫害和其他伤害有关的。JA是植物体内的一种生长调节物质,在调节植物生长发育、光合特性及抗逆反应等方面起着重要的作用,比如种子的萌发,植物的衰老和生长,花粉的孕育,球茎的形成,卷须的缠绕等等。
植物体内存在着多种信号途径,不同的信号途径之间相互作用,从而形成一个精密的调控网络,在植物生长的各个阶段调控,促使植物正常地进行生理活动[22]。WRKY基因可以被SA和JA等植物激素诱导,例如NtWRKY-R1的启动子区域包含两个顺式元件,分别对JA信号和生长素信号作出响应。部分WRKY基因也能调控SA和JA的合成。实验表明,拟南芥WRKY28和WRKY46本身由病原体诱导子快速诱导的,但是它们可以诱导ICS1和PBS3基因表达,而ICS1基因和PBS3基因与SA的积累有关,从而能够将病原体触发的防御基因表达与SA的积累联系起来[23]。
1.3莲中WRKY转录因子简介
莲(Nelumbo nucifera),别称有荷花、芙蕖等,双子叶植物,睡莲科。多年生挺水草本植物。在亚洲,莲作为一种食物已经有7000多年的历史,人们种植莲花是为了获得可食用的根茎、种子和叶子。其芽、花、花药、雄蕊、果实、叶、茎、根茎、根等均被用作治疗癌症、抑郁症、腹泻、心脏病、高血压和失眠的中草药。它的种子寿命长,可存活长达1300年,营养根茎可存活50年以上[24]。因此莲花是一种具有农业、药用、文化和宗教重要性的基础植物。莲的药用价值主要是来源于莲生物碱,国内外许多学者对生物碱及其药理作用进行了许多研究[25]。莲生物碱主要集中在莲叶与莲心中,但是存在于莲叶和莲心中的生物碱种类完全不同,莲生物碱药理活性主要有抗炎、抗氧化、抗衰老、降脂减肥、降压、抑菌、抗心律失常、抗动脉粥样硬化和抗肿瘤等等[26]。研究表明, WRKY基因能调控莲生物碱的合成[27],所以了解莲WRKY基因的功能对莲生物碱的合成意义重大。莲的全基因组测序于2013年完成[24],但是对于莲这一水生生物的WRKY基因的研究较少,具体的功能尚未得知。另外,植物激素能促进莲生物碱的合成[28, 29],而植物中众多WRKY基因可以被激素例如SA和JA所诱导,因此探究莲中WRKY转录因子是否参与调控激素介导的生物碱合成过程是可行的。
本研究以植物莲为研究对象,用SA和JA分别处理0h、3h、6h和24h后得到实验样品。通过RNA-seq技术获得转录组数据,分析转录组数据得到候选WRKY基因的电子表达数据,通过生物信息学相关软件分析基因表达水平以及差异基因的分类,尝试探究WRKY基因在SA和JA介导的信号途径中的作用。这些结果对于我们后期研究WRKY基因功能提供了有力的数据支持。
第2章 比较转录组分析莲WRKY基因家族响应激素SA和JA的表达谱
2.1研究内容与方法
2.1.1 内容
⑴对SA和JA处理后的莲叶片转录组进行总体描述:数据量、表达的总基因数、Q30、总体GC含量、新基因数、基因的表达量、差异基因的分类等;



