CO2浓度升高下对植物碳、氮和水关系的影响:FACE的六个重要结论外文翻译资料
2022-12-22 17:31:37

英语原文共 18 页,剩余内容已隐藏,支付完成后下载完整资料
CO2浓度升高下对植物碳、氮和水关系的影响:FACE的六个重要结论
Andrew D. B. Leakey1,2, Elizabeth A. Ainsworth3,1,2, Carl J. Bernacchi3,1,2,Alistair Rogers4,5,Stephen P. Long1,2,5and Donald R. Ort3,1,2,5,*
11406伊利诺伊大学基因生物学研究所,1206W. Gregory Drive,Urban, H,61801, USA
2伊利诺伊大学植物生物学系,Urban, H,61801, USA
3USDA/ARS, Urban, H,61801, USA
4布鲁克海文国家实验室环境科学系, Upton, NY 11973-5000, USA
5伊利诺伊大学作物科学系, Urban, H,61801, USA
摘要:植物对未来CO2水平的响应的预测首先在持续数天至数周的短期实验中得到表征。然而,随后发现对升高的CO2的长期适应性反应在确定植物和生态系统功能方面非常重要。自由空气CO2富集(FACE)实验是在多个季节评估CO2对植物的影响以及在作物的整个生命周期中对植物的影响的结果。在近20年的时间里,FACE已被用于在完全露天的条件下将植被暴露于高浓度的大气CO2。本综述介绍了从这些实验的长期投资中汲取的一些结论。首先,尽管rubisco活性下调,但CO2升高可以长期刺激光合碳增加和净初级生产。其次,升高的CO2提高了氮的利用效率,第三,降低了叶片和冠层的用水量。第四,升高的CO2通过代谢的转录重编程刺激暗呼吸。第五,升高的CO2不直接刺激C的光合作用,但可以在干旱的时间和地点间接刺激碳的增加。最后,作物物种中CO2升高对产量的刺激远小于预期。虽然许多结论在作物系统中得到了最清晰的证明,但所有结论都对自然系统产生了重要影响。
关键词:气候变化;CO2升高;自由空气二氧化碳富集(FACE);Rubisco
目 录
结论1:CO2升高时尽管会出现光合能力驯化,但碳吸收量会增加 2
结论5:C4植物中CO2升高对碳吸收的刺激是间接的,仅在干旱的情况下发生 13
1 引言
大气CO2浓度在工业革命开始前的至少1000年内稳定在270 lmol·mol-1。从那时起,二氧化碳在全球大气层中不断加速。2009年,CO2为384 lmol·mol-1;比过去2000万年的任何时间都高40%(Pagani et al.,1999; Pearson and Palmer,2000)。到本世纪中叶,CO2预计将超过550 lmol·mol-1,到本世纪末,将超过700 lmol·mol-1 (Prentice et al.,2001)。近年来观测的全球CO2增速明显快于联合国政府间气候变化专门委员会(IPCC)第四次评估报告(AR4)的预期(Dyson,2005; Canadell et al.,2007; Hanson et al.,2008)。这被认为是中国和印度新燃煤电厂快速增加排放的结果,以及发达经济体限制其排放的有效行动。尽管在“京都议定书”下采取了初步措施,但世界似乎走上了可能导致CO2超过IPCC排放情景(A1FI)的道路。因此,自中新世早期(Pearson和Palmer,2000)以来,自然和管理生态系统目前都暴露于陆地植被尚未经历的较高CO2水平,并面临着一个预示着不断增加的不确定后果的未来。
了解植物如何应对CO2的快速变化以及发展其适应能力的知识,是了解全球变化的多种相互作用因素(如干旱,温度,臭氧)对陆地生态系统的全面影响的重要初始步骤。这些生态系统产生的服务依赖于食物,燃料,纤维,清洁空气和淡水。过去三十年中进行的各种封闭研究中关于植物如何应对未来CO2预测水平的研究,已经有了大量知识。随着先进科学的发展和潜在机制的揭示,在现实的露天条件下进行实验的需求愈加明显,最终引导出新技术的诞生—— FreeAir Carbon dioxide Enrichment(FACE)(Lewin et al.,1992,1994; Hendrey and Miglietta,2006)。现在已经有15个主要的FACE实验在世界不同地区的不同生态系统上完全大块复制使用,可以得出许多重要结论。本文介绍了从FACE实验中学到的关于植物碳,氮和水关系影响的六个重要结论,并从这些研究中获得了基础证据。
结论1:CO2升高时尽管会出现光合能力驯化,但碳吸收量会增加
最近FACE研究C3植物对CO2浓度升高的光合响应的第一个结论是尽管光合能力适应,但CO2浓度升高会增强光合碳吸收(A)。光合驯化在测量时通常表现为Rubisco最大羧化速率(Vcmax)和驱动RubP再生的最大电子传递速率(Jmax)的降低(reviewed in Long et al.,2004; Nowak et al.,2004;Ainsworth and Long,2005; Ainsworth and Rogers,2007)。早期针对受控环境中生长在盆栽中的C3植物的研究表明,一些物种光合能力的驯化可能会抵消CO2升高对(A)的促进作用(reviewed in Arp,1991; Stitt,1991; Sage,1994)。但近年来自FACE实验的证据大多表明,高浓度CO2下生长的C3植物,尽管Vcmax和Jmax略有下降,但光饱和的光合碳吸收率(Asat)却得到显著促进(Ainsworth and Rogers,2007)。A的促进程度随着物种和实验条件的变化而变化(Nowak et al.,2004; Ainsworth and Long,2005)
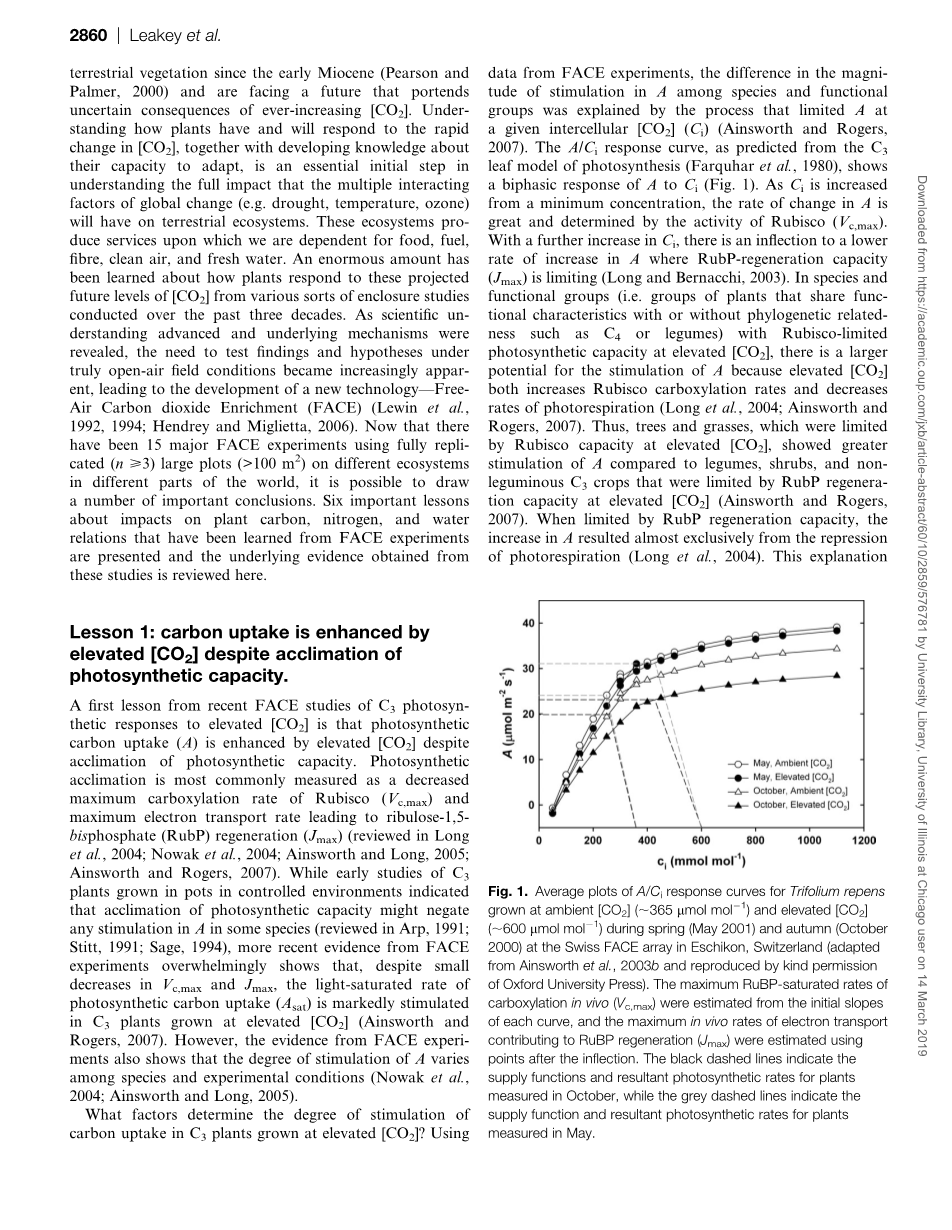
CO2升高对C3植物碳吸收的促进程度受哪些因素决定?运用来自FACE实验的数据,物种和功能组中(A)的刺激幅度的差异通过在给定的胞间CO2浓度(Ci)限制(A)的过程来解释 (Ainsworth and Rogers,2007)。如从光合作用的C3叶片模型预测的A/Ci相应曲线 (Farquhar et al., 1980),显示A至C的双相反应(图1):当Ci从最小浓度增加时,A的增速很大,此时受Rubisco(Vcmax)主导。随着Ci的进一步增加,A增速开始降低,此时RubP再生能力(Jmax)起主导作用(Long and Bernacchi,2003)。在高CO2下光合能力受限于Rubisco的物种和功能群(即具有或不具有系统发育相关性的功能特征的植物群,如C4植物或豆类)中,CO2升高不仅提高了Rubisco的羧化速率,而且降低了光呼吸速率,因此A更易受到促进(Long et al。,2004; Ainsworth and Rogers,2007)。因此,高CO2下受限于Rubisco含量的树木和草类,与高CO2下受限于RubP再生能力的豆科植物,灌木和非豆类C3作物相比,对A的促进作用更大(Ainsworth and Rogers,2007)。当受限于RubP再生能力时,A的增加几乎全来自光呼吸的抑制(Long, 2004)。该解释为在高CO2下树木(46%)和草(37%)中A的促进程度相比平均值的增幅提供了机械基础,相对而言灌木为21%,C3作物为13%,豆类为19%。然而即使在功能群体内,环境和遗传因素也会影响光合作用能力的适应程度和A的刺激。
图1:瑞士埃斯基翁SwissFACE实验
春季(2001年5月)和秋季(2000年10月)生长的环境CO2(365 umol· mol-1)和高CO2(600umol· mol-1)的三叶草的A/Ci响应曲线的平均图。每条曲线的初始斜率可估计体内羧化的最大Rubp饱和速率Vcmax,用映射后的点来估计Rubp再生的电子传递的最大体内速率Jmax。黑虚线表示10月份测量的植物的供应函数和合成光合速率,灰虚线表示5月份测量的。
一般而言,限制沉降强度发展的环境因素,实验因素或遗传因素能使植物更容易出现光合能力驯化,从而在高CO2下减少对A的促进(reviewed in Arp,1991; Stitt,1991;Long,2004;Ainsworth and Rogers,2007)。这一点从不同大小盆中植物在受控环境中生长,其沉降强度会受到生根体积的限制(Arp,1991; Thomas and Strain,1991)这一现象中得到证实。实地研究证实,环境、遗传或管理手段因素使沉降量降低或不足时,会导致叶片碳水化合物增加,随后出现光合能力下调(reviewed in Long et al.,2004; Rogers and Ainsworth,2006; Ainsworth and Rogers,2007)。例如,在管理的黑麦草和三叶草中暴露于高温CO2十年的草地 (Luuml;scher, 2006),季节(Ainsworth, 2003b)和收获实践 (Rogers, 1998; Isopp, 2000; Ainsworth,2003a)在确定L. perenne和T. repens对高温CO2的光合响应中起重要作用。春季,当白天时间长、辐射高时,三叶草草皮生长很快,植物的沉降不太可能受到限制,此时环境CO2和高CO2下植物的光合能力没有差异(图1中的圆符号; Ainsworth, 2003b)。在从A/Ci曲线到y轴的供应函数(图1中的灰色虚线)之后,说明春季在CO2升高后A增加了40%。另一方面,秋季,三叶草受到低温和夜间霜冻的影响,草皮的生长明显比春季少。这些环境条件导致秋季高CO2下Vcmax和Jmax的显著下调(图1中的三角形符号)。10月供应函数(图1中的黑色虚线)显示:秋季CO2升高后A的增幅减半至仅22%(Ainsworth et al., 2003b)。除了改变沉降的环境因素外,在Swiss FACE实验中,定期收获地上组织也改变了源与库的平衡(Luuml;scher等,2006)。在黑麦草的定期收获中去除源组织后,光合能力没有立即变化(Rogers等,1998; Ainsworth等,2003a),但收获后大约3周Vcmax和Jmax的显著减少,此时叶子完全膨胀,且在高CO2下的源容量超过了库容量(Ainsworth等,2003a)。Swiss FACE实验清楚地证明了环境(季节)和实验(收获机制)因素如何改变了源 - 库平衡,光合能力对CO2升高的适应性,以及因此对碳吸收的刺激程度。
遗传因素也可以在光合作用对升高的CO2反应中起重要作用。在Pop FACE实验中暴露于高CO2下的杨树可能最清楚地证明了这一观点(Scarascia-Mugnozza
剩余内容已隐藏,支付完成后下载完整资料
资料编号:[21029],资料为PDF文档或Word文档,PDF文档可免费转换为Word



